アブシシン酸(ABA)について

ABA代謝酵素(CYP707A)における機能解析
気孔閉鎖や種子休眠などのABAの生理作用は内生ABA量の変動によって引き起こされます。ABA量は合成と不活性化のバランスによって制御されていま す。ABAの合成では、9-シス-エポキシカロテノイド・ジオキシゲナーゼ(NCED)が鍵酵素であり、不活性化では、ABA8‘位水酸化酵素が鍵酵素で あると知られています(図2)。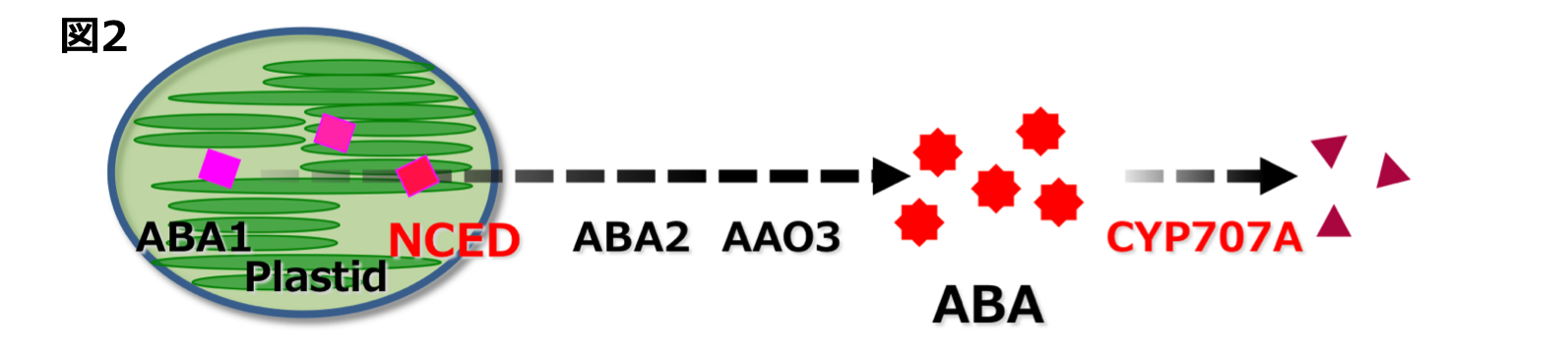 世界で初めてABA8‘位水酸化酵素の実態がCYP707A 遺伝子であることを明らかにして(トロント大:南原先生、明大:久城先生との共同研究)、これまでABA不活性化の分野において様々な知見を明らかにしてきました。CYP707Aの主な働きとして、過剰蓄積したABAを不活性化する(図3A: 農工大:梅澤先生との共同研究)、種子発芽時にはCYP707Aが活性化しABAを分解して発芽を促進している(図3B)、冠水時にCYP707Aが反応しABA量を減少させてシュートの成長に影響を及ぼす(図3C: 名大:中園先生との共同研究)、高い湿度にCYP707Aが応答して気孔開口に関わることなどが明らかとなりました(図3D)。つまり、様々な環境にCYP707Aが応答し、内生ABA量を変化させることで、多様な植物の生理現象を引き起こしていたのです。環境の変化による CYP707Aの応答をさらに詳細に調べることで、植物の生理現象を分子レベルで理解できるのではないかと期待しています。
世界で初めてABA8‘位水酸化酵素の実態がCYP707A 遺伝子であることを明らかにして(トロント大:南原先生、明大:久城先生との共同研究)、これまでABA不活性化の分野において様々な知見を明らかにしてきました。CYP707Aの主な働きとして、過剰蓄積したABAを不活性化する(図3A: 農工大:梅澤先生との共同研究)、種子発芽時にはCYP707Aが活性化しABAを分解して発芽を促進している(図3B)、冠水時にCYP707Aが反応しABA量を減少させてシュートの成長に影響を及ぼす(図3C: 名大:中園先生との共同研究)、高い湿度にCYP707Aが応答して気孔開口に関わることなどが明らかとなりました(図3D)。つまり、様々な環境にCYP707Aが応答し、内生ABA量を変化させることで、多様な植物の生理現象を引き起こしていたのです。環境の変化による CYP707Aの応答をさらに詳細に調べることで、植物の生理現象を分子レベルで理解できるのではないかと期待しています。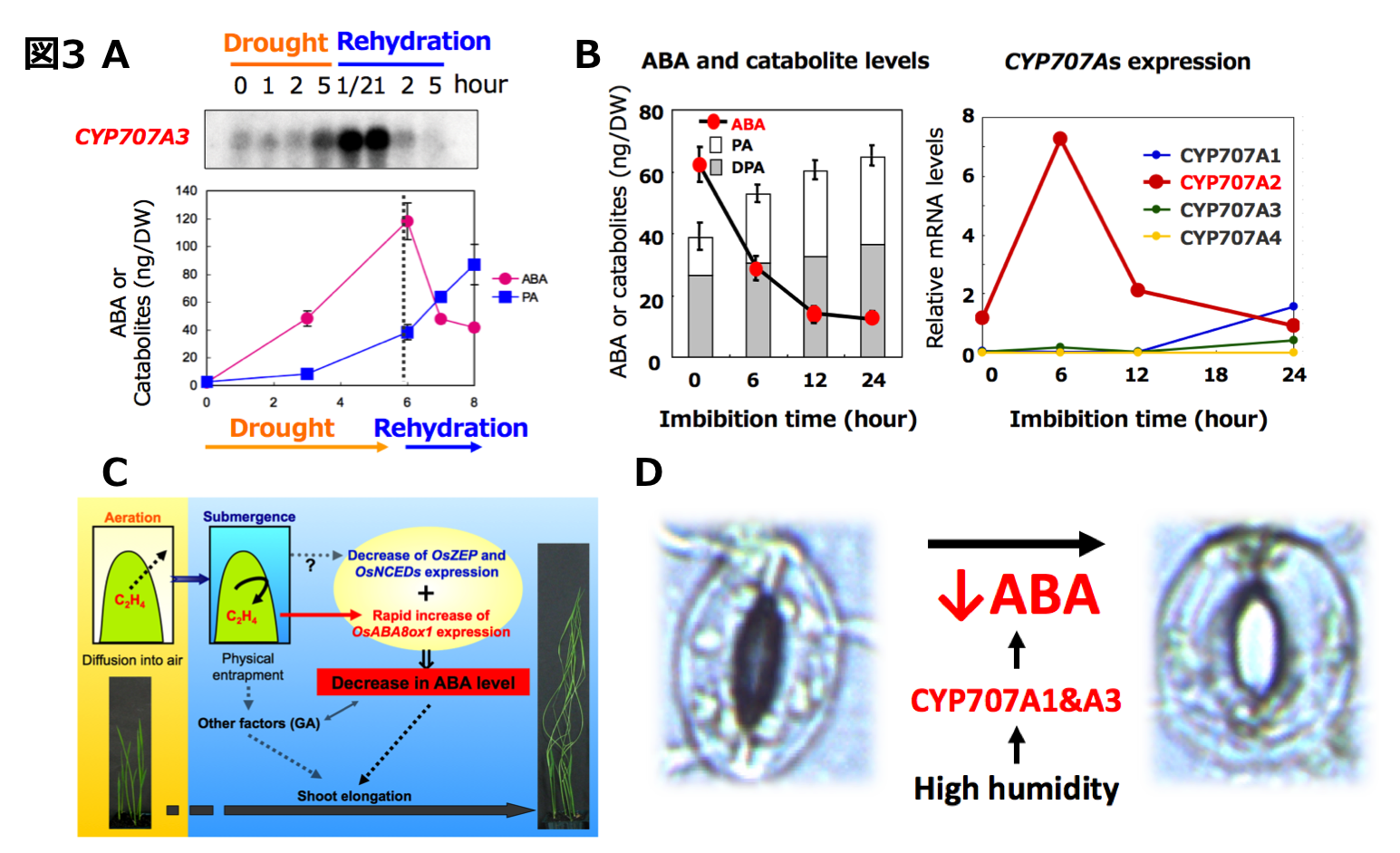
ABA代謝に作用する化合物の発見とその利用
シロイヌナズナのようなモデル植物ではCYP707Aの変異株は簡単に利用できますが、実際の作物では簡単ではありません。ABAの不活性化酵素(CYP707A)がP450酵素であることに着目し、すでに市販されている様々なP450阻害剤が CYP707Aの活性を阻害するかを調べた結果(神戸大の水谷先生との共同研究)、現在よく使われているウニコナゾールという農薬がCYP707A活性を強く阻害することを発見しました。実際に、ウニコナゾールを植物に投与すると、ABA量が増加し、葉からの蒸散量の低下や乾燥耐性の向上が見られました。既に市販されている阻害剤の新たな利用価値を発見できたことは農学的に意味があります。しかし、ウニコナゾールは、元々ジベレリンという成長促進に関わる植物ホルモンの阻害剤であることから、矮化も引き起こしてしまいます。そこで、CYP707A特異的な阻害剤の開発を静岡大の轟先生と竹内先生らの研究グループと行い、矮性を引き起こさない乾燥耐性を示すアブシナゾール-E3Mの開発に成功しました(図4)。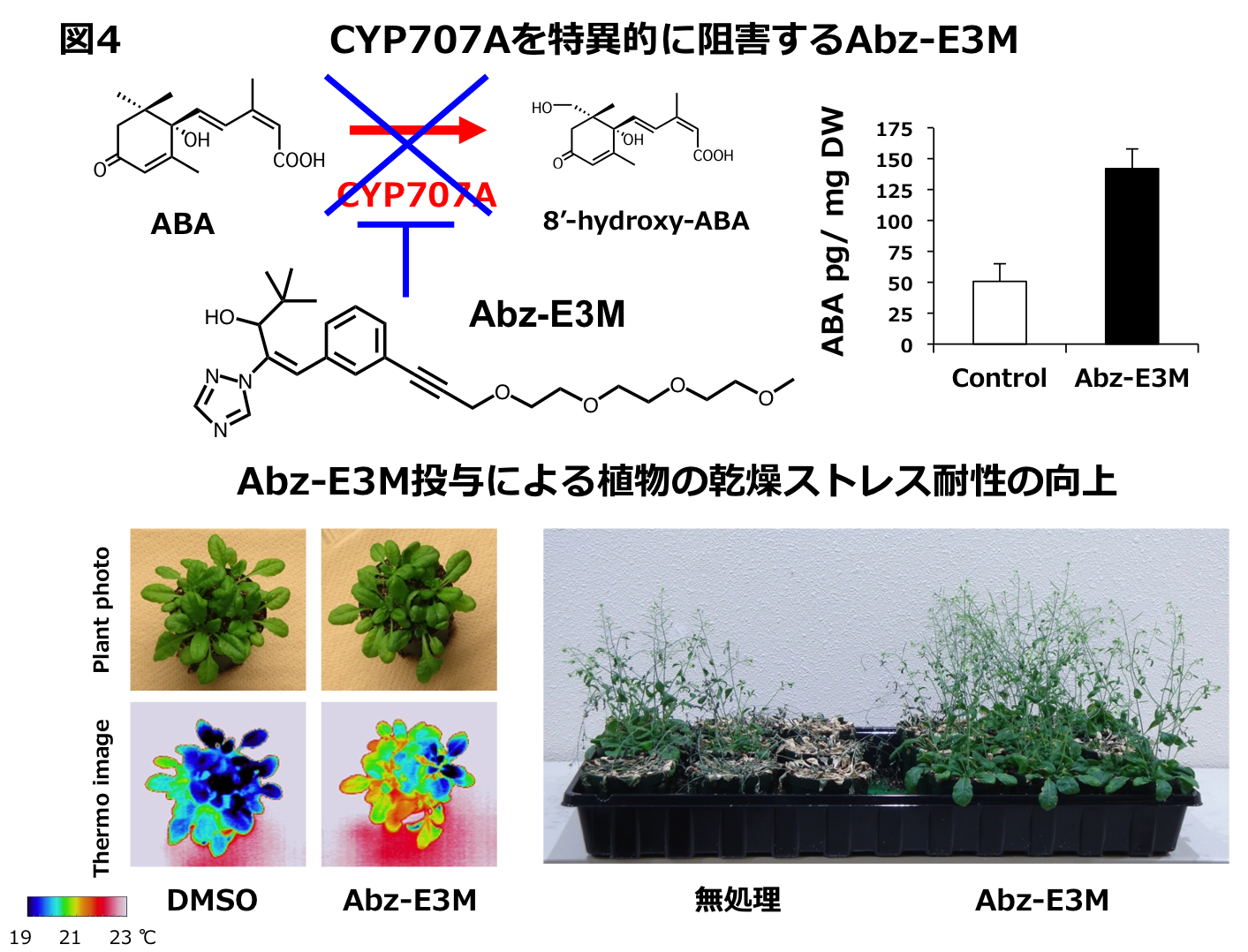
ABA受容体に作用する化合物の探索とその応用
ABAが植物の乾燥耐性を向上する一方で、低濃度のABAを通常のフィールドで投与すると、植物の生長を促進させることが知られています。したがって、様々なABAの農業的応用が考えられます。しかし、天然あるいは人工合成のABAの価格が高価であること、ABAが光に弱いこと、投与しても植物体内で不活性化されてしまう等などの問題があるため、現在のところ、市場ではABAの利用と応用がそれほど普及していません。一方、ABAとは構造の全く異なる合成化合物でABA受容体に結合できる化合物(アゴニスト)としてピラバクチンが報告されていました。しかし、ピラバクチンは活性が弱いために、種子の発芽阻害は引き起こすものの、植物の乾燥耐性を向上させることができません。カリフォルニア大学リバーサイド校のCutler教授との共同研究で、乾燥ストレスを寄与する新奇のABAアゴニストとしてキナバクチンを発見しました(図5A)。さらに、乾燥ストレス耐性や種子発芽阻害を示さないユニークな新奇ABAアゴニストを開発し(図5B)、これらを用いたケミカルゲノミクスによって、ABA受容体の生理的機能の解明やABA感受性の変化した新奇変異体の単離も進めています。一方で、ABAの作用を弱らせる化合物(アンタゴニスト)の開発を静岡大学の轟先生と竹内先生と共同で行っています(図5C)。ABAアンタゴニストは、ABAの生理作用を理解するためのバイオプローブとして利用できるだけでなく、農場上問題となる過剰なABAの作用を、適切にコントロールできる新しい植物成長調節剤としても利用できることが期待できます。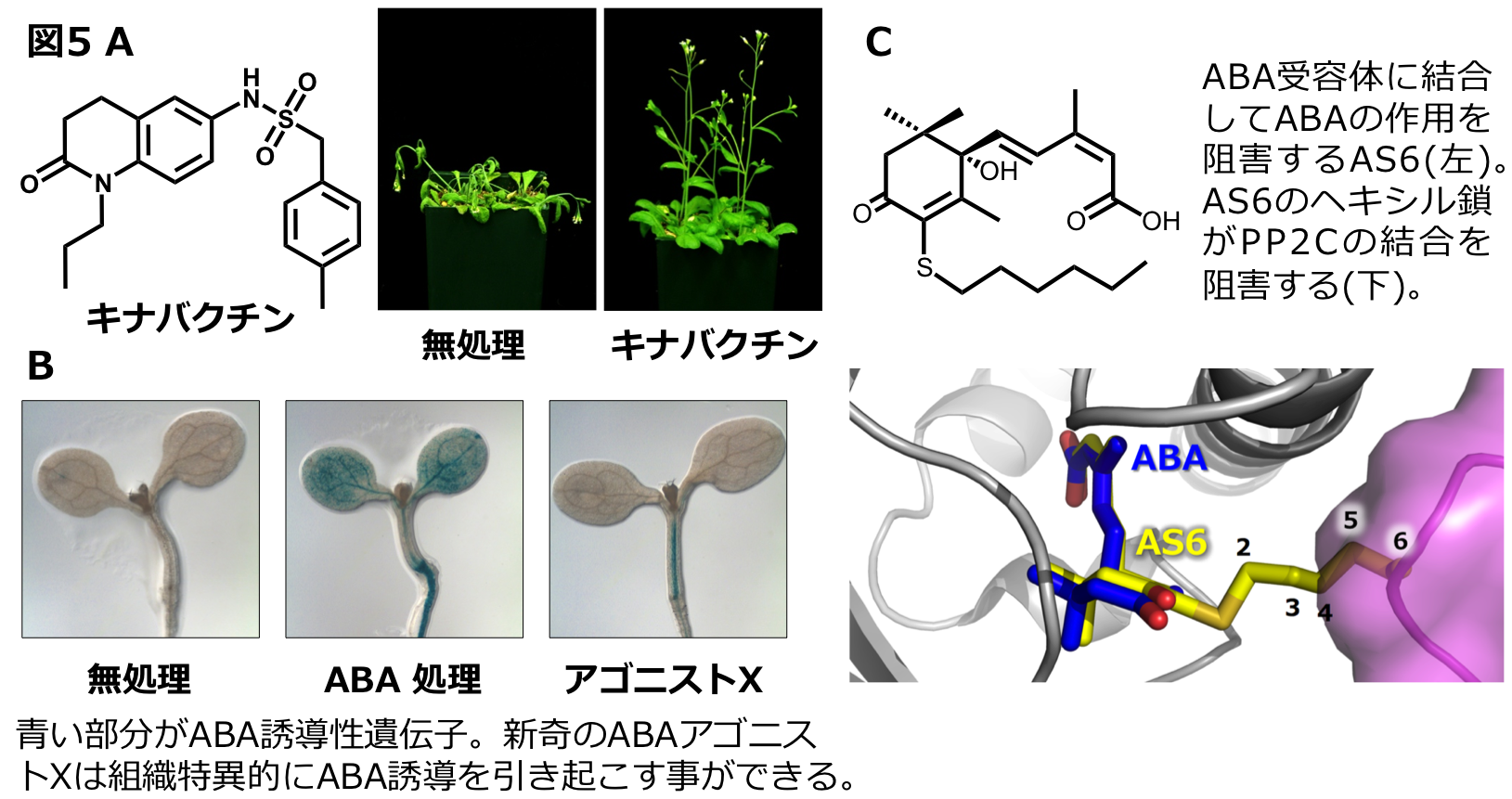
ABAの利点を生かした作物研究
CYP707Aの機能を破壊した変異株やABAの感受性を向上した植物では、種子休眠性と乾耐性の向上が見られます(図6A)。 これらの実験結果は、モデル植物のシロイヌナズナで得られたものですが、他の植物種でもABAの作用を向上させることで、同様の形質がもたらされるのではないかと考えています。ABAの感受性が向上したコムギを創出し、水が貴重な乾燥地域でも作物生産が可能な節水性耐乾性コムギを遺伝子組換え技術により創出し、その形質を調べています(図6B: 鳥取大学の辻本先生と妻鹿先生、および作物研究所の安倍先生との共同研究)。そして、これらの知見を応用し、遺伝子組換えに頼らない方法として、約2万の多様な遺伝的形質を持つコムギ系統からケミカルスクリーニングによって耐乾性コムギ選抜し、形質を解析しています。単離した節水型耐乾性コムギ系統は、鳥取大学乾燥地研究センターと国際乾燥地農業研究センター (ICARDA)との共同研究により、海外の乾燥地にて、収量評価実験を計画しています。選抜した耐乾性コムギが、世界のコムギの耐乾性を向上させることのできる新しい育種素材になり、最終的には、世界の食糧生産の向上に貢献できることを願って研究しています。